Annals of
Burns and Fire Disasters - vol. XIII - n. 3 - September 2000
ANALYSIS OF
PROLIFERATION/DIFFERENTIATION AND IMMUNOGENICITY
OF CULTURED HUMAN KERATINOCYTES AND NORMAL HUMAN EPIDERMIS
Garcia Fernândez E.,1 Maruri N.,1 Arrieta A.,1 Ririôn M.,1 Arranz M.C.,1 Bejar J.M.,2
Garcia Masdevall M.D.,1 Gabilondo F.J.2
1Department of Immunology,Cruces
Hospital,Baracaldo,Vizcaya, Basque Health Service, Spain
2Plastic Surgery and Burns, Cruces Hospital
SUMMARY. Normal
human keratinocytes can be serially cultured in vitro and under appropriate culture
conditions to give epidermal sheets which may be used to cover deep and large skin defects
or burns. The analysis by flow cytometry of epidermal cells in normal human epidermis and
cultured human keratinocytes is proposed to predict the optimal time for the grafting of
in vitro prepared allogeneic keratinocytes. This analysis regarded their proliferative and
immunogenic stage, and the evaluation of antigens whose level of expression is related to
these. In 19 skin biopsies and 22 cultured epidermal sheets we analysed: markers of
epidermal proliferation/differentiation (31 integrins (CD29) and Kl/K10 intracytoplasmatic
keratins; antibodies of immunogenicity anti-HLA-DR and anti-HLA A,B,C, and antibodies
which identify other epidermal cells (anti-vimentin and anti-CD45. We made the following
observations. First, the phenotype of epidermal cells that were obtained from cultured
epidermal sheets was similar to the phenotype of isolated cells from normal human skin in
samples treated with lysolecithin. Second, the CD29+ cells increased compared with the
CD29-K1/K10+ in normal human skin and cultured sheets, especially the CD29+K1/K10+ group,
which was suprabasal but occasionally basal and highly proliferative. This could be due to
the high proliferative ability of keratinocytes in culture. Third, the cultured
keratinocytes lost nearly all expression of the HLA class II and class I antigens. The
cultured epithelial sheets in the laboratory can therefore be grafted successfully and we
may suppose that immunological rejection does not occur.
Introduction
The degree of
maturation and the metabolic stage of epidermal cultured sheets prior to grafting is
important because there is a relationship between graft survival and its metabolic
stage.The ability of keratinocytes to maintain proliferative potential after seeding and
their ability to form a properly oriented epithelium in vitro would suggest that
autologous keratinocyte grafting may be suitable as a wound dressing
The epidermis has traditionally been divided into three functionally different layers:
germinative, differentiated, and cornified. Some evidence suggests that the germinative
compartment is heterogeneous. Two groups of antigens are closely related to the
microanatomic location and the stage of cellular differentiation in the epidermis:
adhesion molecules and keratins.The determination of (32 integrins (CD29) and KI/K10
intracytoplasmic differentiation keratins by flow cytometry, in combination with optical
light scatter characteristics of the cells, identifies the proliferative compartments of
in vivo human epidermis.
Two different proliferating cell types have been described in human epidermis.One subset
of normal epidermis, predominantly basal, is composed of slow cycling, small cells with a
primitive cytoplasmic organization (CD29+ Kl/K10-). The other proliferative subset of
normal epidermis, which is suprabasal and occasionally basal, is highly proliferative and
larger in size, and exhibits a more complex cytoplasmic structure (CD29+ K1/K10+). The
cells that express only the K1/K10 marker are the differentiated keratinocytes of skin
(CD29K1/K10+).
Human epidermal cells have been successfully grown in serial culture to form keratinizing
colonies. The most satisfactory technique, which is based on those of Rheinwald and Green
and of Green, uses serum-containing medium and a feeder layer of lethally irradiated
murine 3T3 fibroblasts which is required to initiate formation of the keratinocyte colony.
Additional supplements enhance the proliferation, increase the life span of the
keratinocyte cultures, and permit a higher proportion of small proliferating cells. The
study of (32 integrins Kl/K10 keratins allows the proliferative stages of cultured human
keratinocytes to be distinguished.
Although expression-of Kl/K10 is specific for keratinocytes, the anti-CD 29 recognizes the
(32 integrin chain and may be expressed on other cells in the epidermis such as
melanocytes, Langerhans cells, or infiltrating leukocyte populations. Although the number
of these cells in normal epidermis is relatively low compared with that of the
keratinocytes, their staining has been realized in order to determine their contribution.
Anti-vimentin, anti-HLADR, and anti-HLA A,B,C antibodies have been used to identify them.
Since keratins K1/K10 and vimentin are intracytoplasmatic, it is necessary to make the
cells permeable to these antibodies and to allow the appropriate dye molecules to reach
the respective antigens in the cell interior. At the same time, the antigens must be
preserved in their natural, antigenic conformation, and leakage out of the cell must be
prevented. Several protocols for cell preparation and staining are generally applicable,
and thus the cells become permeable to lysolecithin and ethanol. Ethanol dramatically
alters the cell morphology, cell aggregation is increased, the antigens lose their
distribution and antigenicity, and the background staining is usually more apparent. In
view of these facts we used lysolecithin.
Lysolecithin (lysophosphatidylcholine) is a lipid with potent membrane-active
characteristics. Lysolecithin replaces phospholipids in the membrane bilayer with a
resulting loss of integrity of the membrane, thereby allowing the passage of molecules the
size of IgG (=150 kDa). At lower concentrations, lysolecithin can serve to make the cell
surface membrane permeable without complete lysis of the cell.
The study and quantification of Class 1 and 11 HLA antigens in epidermal cultured sheets
enable us to know at what moment epidermal cells diminish their capacity to initiate an
allogeneic immune response.
The study of anti-HLA-DR, present in the Langerhans cells, and of anti-HLA A,B,C, present
in all epidermal cells at different levels, enables us to evaluate the immunogenicity of
epidermal cells extracted from skin biopsies and cultured sheets. Some studies provide
evidence that when human epidermal cells are grown in culture, they lose their expression
of HLA-DR antigens, but the cells used in organ equivalent grafts express class I MHC
antigens.
Materials
and methods
Epidermal cell suspension
The 41 skin samples - biopsies taken from
normal skin and split-thickness skin grafts from burns - were placed in culture medium and
transferred to the laboratory. After removal of as much subcutaneous tissue and dermis as
possible, the tissue was cut into smaller fragments. These were then digested with 0.17%
trypsin solution (Seromed. Biochrom KG) at 4 °C overnight, followed by treatment with
trypsin-EDTA (25% - 0.02 mM) solution Seromed. Bliochrom KG) for 10-20 min to ensure
dissociation into single cells 2' These cells were studied by flow cytometry or seeded in
plastic flasks to obtain epidermal sheets.
Keratinocyte culture method
Plastic tissue culture flasks of 75 cm2
(Costar Co.) already containing 2.5 x 106 lethally irradiated murine 3T3 fibroblasts (European
Collection of Animal Cell.Cultures) were inoculated with over 3 x 104
epithelial cells/cm2. The cultures were fed with a 3:1 mixture of the
Dulbecco-Vogt modification of Eagle's medium and Ham's F-12 medium (Gibco BRL Co.) with
supplements described by Rheinwald and Green. The cultures were incubated at 37 °C in a
humid 95% air/5% COZ environment (Selecta Co.) and the medium was changed every three
days.
To prepare secondary cultures,
subconfluent primary cultures were trypsinized and the cells transferred to flasks the
same size containing irradiated 3T3 cells. These cultures were maintained in the same way
as the primary cultures until they reached a confluent layer of keratinocytes.
For the cytometry flow study, the cells were detached with trypsin solution from 75 cm2
plastic flasks and resuspended in, culture medium.
Cell permeabilizing method
Epidermal cells were
isolated from 19 skin biopsies and the cultured keratinocytes were obtained by seeding
epidermal cells from 22 samples.
The cells were permeabilized using lysolecithin (lysophosphatidylcholine) (Sigma Chemical
Co.). This treatment was performed in phosphate-buffered saline (PBS) pH 7.2 without
calcium or magnesium (Gibco BRL Co.), at a concentration of 1-5 x 106
cells/ml. Lysolecithin was stored in aliquots at -20 °C at a stock concentration of 20
mg/ml in absolute methanol (Merck KGaA). Immediately prior to use, an aliquot was brought
to room temperature, diluted in cold PBS, and added to the cells to yield a final
concentration of 50 ~tg of lysolecithin per ml. This mixture was incubated at 4 °C for 5
min, diluted at least twofold in PBS containing 1% bovine serum albumin (BSA) (Merck KGaA)
and 0.02% sodium azide (Merck KGaA). The cells were then washed with this solution.
Staining procedure and flow cytometry
analysis Cell suspensions were stained using direct or indirect immunofluorescence
staining procedures. The mAbs used were:
- anti-CD29 (Coulter Immunology) and anti-K1/K10 (Cymbus
Bioscience Ltd), markers of epidermal proliferation/differentiation;
- antibodies of immunogenicity anti-HLA-DR (Becton Dickinson
Co.) and W6/32 anti-HLA A,B,C conformational (American Type Culture Collection, Rochville,
MD);
- antibodies which identify the other epidermal cells:
anti-vimentin (Cymbus Bioscience Ltd) and antiCD45 (Becton Dickinson Co.), using FITC-Goat
F(ab') 2 mouse anti-IgG (H+L) human antibody (Becton
Dickinson Co.) for indirect techniques. Flow cytometry was performed using a FACS SCAN
(Becton Dickinson Co.).
Results
Identification and quantification of
the different compartments in skin and cultures
The cell population obtained was very
heterogeneous in size and complexity and the separation of cells into groups would be
somewhat arbitrary. Lysolecithin-permeabilized cells had a similar distribution to those
that were not permeabilized, except for small-size cells, because treatment with
lysolecithin resulted in partial disruption of the plasma membrane, with a resulting loss
of cytoplasm and a decrease in the intensity of forward light scatter. However, the
nucleus and cytoplasmic structures such as mitochondria, membrane vesicles, and
intermediate filaments apparently remained intact, so that cells retained their
distinctive differences in size light scatter.
In normal human epidermis, all the DR+, CD45+, and vimentin+ cells showed double staining
with anti-CD29, indicating that (31 integrin is indeed present in Langerhans cells,
leukocytes, and melanocytes. The number of these cells in the normal epidermis is
relatively low compared with the keratinocytes, which comprise about 90% of epidermal
cells. The cell percentages that express each marker as the percentage of total epidermal
cells from biopsies (Table I) are similar to the percentages described by others.
| |
|
CD 29+ |
CD 45+ |
HLA DR+ |
Vimentin+ |
K1/K10+ |
| |
|
Biops. |
Sheets |
Biops |
Sheets |
Biops |
Sheets |
Biops |
Sheets |
Biops |
Sheets |
| |
x (%) |
|
|
|
|
|
Lysolecithin
permeabilized |
on-1 |
|
|
|
|
|
Cells |
n |
|
|
|
|
|
|
Table I -
Percentage of cells expressing each marker as a percentage of total isolated epidermal
cells and cultured cells (photype of isolated epidermal cell populations) |
|
Identification and quantification of basal and suprabasal cell compartments in normal
epidermis and epidermal sheets
The evaluation of cell compartments and markers must be done using CD45- cells. The
proliferative compartment of human in vivo epidermis contains two subpopulations
characterized by CD29 and Kl/K10 expression! CD29 identifies all basal keratinocytes in
normal skin, so that the predominantly basal cells were defined as CD29+K1/K10, implying
proliferative ability. We observed in studied samples from biopsies and sheets that the
percentage of CD29+ cells (Fig. 1) was close to the results of other researchers, although
slightly higher.
The cytoplasmic expression of the differentiation keratin pair, KI/K10, was used to
separate more highly differentiated cells from the proliferative compartment cells. The
appearance of K1/K10 keratins in keratinocytes is considered to be one of the first signs
of differentiation.The suprabasal cells were identified on the basis of their positive
expression of K1/K10, and they could be divided into two populations depending on their
coexpression of CD29. Undifferentiated CD29+ K1/K10cells in the basal layer express K1/K10
upon commitment to differentiation and separation from the basement membrane. The increase
in the CD29+ group is due to the CD29+ K1/K10+ cells which increased in biopsies and
cultured cells; the CD29- K1/K10+ cells decreased compared with the results of other
researchers! The percentage of CD29+ Kl/K10+ cells in biopsies and
lysolecithin-permeabilized sheets was consistent and slightly higher than that found by
other researchers.
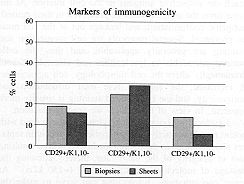
|
Fig. 1 - Quantification in percentages of cellular
compartments in normal epidermis and epidermal
sheets: basal (CD29+ Kl/K10-), suprabasal (CD29+ Kl/K10+), and differentiated (CD29-
Kl/K10+) cells. |
|
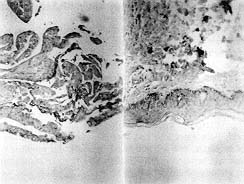
|
Fig. 2 - Histological
sectioning: a) adult skin and b) secondary culture at least 30 days old. Note the basal
layer marked CD29+ and the keratohyaline granules in intermediate layer cells and the most
superficial cells. |
|
Fig. 2 shows a secondary culture at least 30 days old
which attained thicknesses ranging between four and six cell layers. During the latter stage of growth, secondary cultures were composed
of small, rounded basal cells attached to a plastic substrate and suprabasal layers
consisting of enlarged, irregularly shaped vacuolated cells resembling the stratum
germinativum and spinosum. Although no stratum corneum developed in these cultures, the
cells retained the capacity, once grafted, to differentiate terminally. The basal layer
was also more evident and organized (CD29+), and the differentiation appeared in the
expressivity of intermediate and superficial layers (Kl/K10+). Occasionally a few
keratohyaline granules in cells of the intermediate layer and in the most superficial
cells were found.
Immunogenicity of epidermal sheets
Immunological reactivity is generally linked to
the presence of class I and class II MHC antigens on the surface of cells. In our work the
expression of HLA class II antigens in epidermal cells (4.98 ± 3.51%) was similar to that
of normal skin cells (4.39 ± 1.63%) (Table II) and both were weakly immunogenic. The HLA
class I antigens, present in all epidermal cells (88.23 ± 5,08%) showed almost total loss
of their expression in the cultures (4.94 ± 3.22%) (Table II). To determine these results
with regard to HLA-DR and HLA A, -B markers, CD45- cells were used.
| |
HLA DR+ CD 29+ |
HLA A, B, C+ |
| |
Biopsies |
Sheets |
Biopsies |
Sheets |
% |
|
|
on-1 |
|
|
n |
|
|
|
Table II -
Percentages of class I and class H MHC antigens on the surface of normal skin cells and
epidermal culture cells. Phenotype of isolated epidermal cell population. |
|
Cultured cells have similar phenotypes
to the epidermal cells of human skin and the cultures could therefore have similar
behaviour to that of adult skin. The low values of K1/K10+ cells were consistent with
histological and immunohistochemical results, in which cultured epithelial sheets showed a
similar structure to normal epidermis although the state of differentiation was not
complete. This decrease of K1/K10+ cells was also observed when the basal and suprabasal
cells compartments were analysed, especially in the CD29- K1/K10+ group in epidermal
sheets. The granulous layer was absent and the superficial cells showed a lower level of
keratinization.26 This partially differentiated sheet retained its capacity to
differentiate further when grafted onto the wound and could be used as an autograft. This
is very important, for establishing a mechanical barrier and for linking the grafted sheet
to the underlying connective tissue.
The high percentage of CD29+ cells observed could be due to the use of samples from young
donors in the skin biopsies2b because the growth potential of basal keratinocytes declines
as the age of the donor increases. In culture the use of biopsies from young donors
provides a high percentage of CD29+ cells ,because colony-forming efficiency, which is an
expression of the basal cells' capacity to grow, declines with the patients' increasing
age. Cell passages in vitro and natural aging in vivo cause a progressive decrease in
cells that possess the highest growth potential (holoclones or CD29+ cells) and a
progressive increase in cells that possess the lowest growth potential and with a lifetime
of no more than 15 generations (paraclones or CD29- cells.
It is important to note that epithelial cells in culture almost entirely lose their
immunological ability because of the expression of HLA Class I antigens. At least four
cell types in the epidermis may contribute to the immunogenicity of skin:" the
Langerhans cells, the thymus-derived dendritic cell, the melanocyte, and the keratinocyte.
Some results have shown that human epidermal Langerhans cells also undergo profound
morphological and phenotypic changes during in vitro culture of epidermal cells. Some
investigators" have observed the Langerhans cell phenotype as early as 24' to 48 h
after isolation of epidermal cells and that their class II and class 1 antigenic density
dramatically increases.This is important because in the skin initiation of the immune
response has been primarily attributed to class II antigen-bearing Langerhans cells and to
subsets of immunogenic cells in the dermis (macrophages, lymphocytes, and capillary
endothelial cells). The Langerhans cells are probably critical cells for skin graft
rejection when skin from one individual is transplanted to another," but the
successful grafting of cells across major histocompatibility barriers suggests that
grafted cells are either non-immunogenic or so weakly immunogenic that immunological
rejection could not be detected." The failure of cultured keratinocytes to be
rejected could be associated with the loss of cells expressing class II antigen during
culture. It has been demonstrated that cultured epidermal cells are devoid of Langerhans
cells and other class II MHC-bearing cells.
Melanocytes grow in the same culture conditions that allow keratinocyte growth, and their
growth is specifically induced by epithelial cells. For this reason melanocytes are the
most frequently found cells in epidermal cellular suspension;' however, the expression of
MHC class I and II antigens has been shown to be deficient in these cells and in
Langerhans cells." This may be confirmed by our study.
Keratinocytes and dermal fibroblasts do not consistently express class II antigens"
but there are several diseases in which this happens (graft-versus-host disease, mycosis
fungoides, allergic contact dermatitis, lichen planus, tuberculoid leprosy)." Morhenn
et al." have shown that interferon can induce this keratinocyte class II synthesis.
The role of keratinocyte surface class Il in immune responses is not known but Roberts et
al." have suggested that keratinocyte class II expression may regulate Langerhans
cell traffic in the epidermis.
In view of our current results, with cultured keratinocytes almost entirely losing their
immunological capacity, we might suppose that the immunological rejection did not exist.
Although several investigators, using a variety of techniques, have failed to demonstrate
the survival of cultured keratinocyte allografts in humans, it does not appear that the
grafts are rejected, but instead that they are gradually replaced by recipient cells. The
complex immunological response to cultured keratinocyte allografts has not been fully
elucidated and continues to be an active area of research, and we still need to study the
problem of cultured keratinocyte survival in vivo if these results are to be fully
confirned.
RESUME. Il est possible de cultiver les
kératinocytes humains normaux en série et dans les conditions culturelles appropriées
pour produire des lames épidermiques pour couvrir les défauts profonds et étendus de la
peau ou les brűlures. Les Auteurs proposent la cytométrie du flux des cellules
épidermiques dans l'épiderme humain normal et les kératinocytes humains cultivés pour
prédire le temps optimal pour le greffage des kératinocytes allogčnes préparés in
vitro. Cette analyse concernait leur phase proliférative et immunogčne et l'évaluation
des antigčnes dont le niveau d'expression est corrélé avec eux. Dams 19 biopsies
cutanées et 22 lames épidermiques cultivées les Auteurs ont analysé: les marqueurs des
intégrines (CD29) (31 de la prolifération/différentiation épidermique et des
kératines intracytoplasmatiques Kl/K10; les anticorps de l'immunogénicité anti-HLA-DR
et anti-HLA A,B,C, et les anticorps qui identifient d'autres cellules épidermiques
(anti-vimentine et anti-CD45). Les Auteurs ont fait les observations suivantes:
premičrement, le phénotype des cellules épidermiques obtenues des lames épidermiques
cultivées est semblable au phénotype des cellules isolées de la peau humaine normale
dans les échantillons traités avec la lysolécithine; deuxičment, les cellules CD+29
sont augmentées par rapport au CD29-Kl/10+ de la peau normale et les lames cultivées,
particulierčment dans le groupe CD29+K1/K10+, qui est suprabasal mais quelquefois basal
et extręmement prolifératif (ceci pourrait ętre causé par la capacité proliférative
élévée des kératinocytes cultivés); troisičmement, les kératinocytes cultivés
perdent presque toute l'expression des antigčnes HLA de classe I e II. Les Auteurs
concluent que les lames épithéliales cultivées en laboratoire pourront ętre greffées
avec succčs et qu'il est possible que le rejet immunologique ne se manifeste pas.
BIBLIOGRAPHY
Staiano-Coico L., Higgins
P.J., Darynkiewicz Z. et al.: Human keratinocyte culture. Identification and stating of
epidermal cell subpopulations. J. Clin. Invest., 77: 396-404, 1986.
Franzi A.T., D'Anna F., Zicca A., Trabucchi
E.: Histological evaluation of human cultured epithelium before and after grafting. Burns,
18: 26-31, 1992.
Clausen OTT., Potten C.S.: Heterogeneity of
keratinocytes in the epidermal basal cell layer. J. Cutaneous Pathol., 17: 129, 1990.
Pavlovitch J.H., Rizk-Rabin M., Jaffray P.,
Hoehn H., Poot M.: Characteristics of homogeneously small keratinocytes from newborn rat
skin: Possible epidermal stem cells. Am. J. Physiol., 261: C964, 1991.
Staquet M.J., Dezutter-Dambuyant C.,
Zambrung G., Schmitt D.: Human epidermal basal keratinocytes express CDw29 antigens. Br.
f. Dermatol., 121: 577-85, 1989.
Fleischmajer R., McDonald E.D., Contard P.,
Perlish J.S., Immuno chemistry of a keratinocyte-fibroblast co-culture model for
reconstruction of human skin. J. Histochem. Cytochem., 41: 1359-66, 1993.
Galvin S., Loomis C., Manabe M., Dhouailly
D., Sun T.T.: The major pathways of keratinocyte differentiation as defined by keratin
expression: An overview. Adv. Dermatol., 4: 277-300, 1989.
Bata-Csorgo Z.S., Hammerberg C., Voorhees
J.J., Cooper K.D.: Flow cytometric identification of proliferative subpopulations within
normal human epidermis and the localization of the primary hyperproliferative population
in psoriasis. J. Exp. Med., 178: 1271-81, 1993.
Lavker R.M., Sun T.-T.: Heterogeneity in
epidermal basal kerati nocytes: Morphological and functional correlations. Science,
215:1239, 1982.
Lavker R.M., Sun T-T.: Epidermal stem
cells. J. Invest. Dermatol., 81: 121, 1983.
Rheinwald J.G., Green H.: Serial
cultivation of strains of human epidermal keratinocytes: The formation of keratinizing
colonies from single cells. Cell, 6: 331-44, 1975.
Rheinwald J.G., Green H.: Epidermal growth
factor and multiplication of cultured human epidermal keratinocytes. Nature,
265:421-4,1977.
Green H.: Cyclic AMP in relation to
proliferation of the epidermal cell: A new view. Cell, 15: 801-11, 1978.
Pan Si S., Tsou H.C., Lee X., Peacocke M.:
Cultured human melanocytes express the intermediate filament vimentin. J. Invest.
Dermatol., 101: 383-6, 1993.
Morhenn V.B., Benike C.J., Cox A.J.,
Charron D.J., Engleman E.G.: Cultured human epidermal cells do not synthesize HLA-DR. 34.
J. Invest. Dermatol., 78: 32-7, 1982.
Hefton J.M., Amberson J.B., Biozes D.G.,
Weksler M.E.: Loss of HLA-DR expression by human epidermal cell after growth in culture.
J. Invest. Dermatol., 83: 48-50, 1984.
Larsen J.: Measurement of cytoplasmic and
nuclear antigens. In: "Flow cytometry. A practical approach", Ormerod M.G.
(ed.), 2nd edition, Oxford University Press, 106-7, 1994.
Schroff R.W., Bucana C.D., Klein R.A.,
Farrell M.M., Morgan A.C.: Detection of intracytoplasmic antigens by flow cytometry. J.
Immunol. Method, 70: 167-77, 1984.
Bell E., Rosenberg M.: The commercial use
of cultivated human cells. Transplant. Proceed., 22: 971-4, 1990.
Stingl G.: Dendritic cells of skin.
Dermatol. Clinics, 8: 673-9, 1990.
Garcia Fernandez E., Bejar J.M.,
Alonso-Varona A., Garcia Masdevall M.D., Gabilondo F.J.: Comparative study of the human
keratinocyte isolation methods and in vitro culture techniques in a single laboratory.
Eur. J. Plastic Surgery, 21: 353-7, 1998.
Jones P.H., Bishop L.A., Watt F.M.:
Functional significance of CD9 association with beta 1 integrins in human epidermal
keratinocytes. Cell Adhesion Comm., 484-5: 297-305, 1996.
Moll I., Houdek P., Schmidt H., Moll R.:
Characterization of epidermal wound healing in a human skin organ culture model:
Acceleration by transplanted keratinocytes. J. Invest. Dermatol., 111: 251-8, 1998.
Paramio J.M., Casanov M.L., Segrelles C.,
Mittnacht S., Lane E.B., Jorcano J.L.: Modulation of cell proliferation by cytokeratins
K10 42. and K16. Mol. Cell Biol., 19: 3086-94, 1999.
Schoop V.M., Mirancea N., Fusenig N.E.:
Epidermal organization and differentiation of HaCaT keratinocytes in organotypic
co-culture with human dermal fibroblasts. J. Invest. Dermatol., 112: 34353, 1999.
Garcia Fernandez E., Bejar J.M., Maruri N.,
Garcia Masdevall M.D., Camarero C., Gabilondo F.J.: Histological and immunohistochemical
evaluation of human cultured epidermal cells. Ann. Burns and Fire Disasters, 10: 40-4,
1997.
Jakic-Razumovic J., Zekusic M.,
Vladovic-Re1ja T., Botanic M.: Organotypic skin cultures: A human model for basic studies.
Croatian Med. J., 39: 401-3, 1998.
De Luca M., Albanese E., Bondanza S. et
al.: Multicentre experience in the treatment of burns with autologous and allogenic
cultured epithelium fresh or preserved in a frozen state. Burns, 15: 303-9, 1989.
Michel M., L'Heureux N., Auger F.A.,
Germain L.: From new-born to adult: Phenotypic and functional properties of skin
equivalent and human skin as a function of donor age. J. Cell Physiology, 171: 17989,
1997.
De Luca M., Cancedda R.: Culture of human
epithelium. Burns, 18: 5-10, 1992.
Hansbrough J.F.: Culture epithelium for
wound culture In: "Wound coverage with biologic dressings and cultured skin
substitutes", R.G. Landes Company, Georgetown, Austin, 63-92, 1992.
Romani N., Lenz A., Glassel H. et al.:
Cultured human Langerhans cells resemble lymphoid dendritic cells in phenotype and
function. J. Invest. Dermatol., 93: 600-9, 1989.
Teunissen M.B.M., Wormmeester J., Krieg
S.R. et al.: Human epidermal Langerhans cells undergo profound morphologic and
phenotypical changes during in vitro culture. J. Invest. Dermatol, 94: 166-73, 1990.
Schuler G., Steinman R.M.: Murine epidermal
Langerhans cells mature into potent immunostimulatory dendrific cells in vitro. J. Exp.
Med., 161: 526-46, 1985.
Witmer-Pack M.D., Valinsky J., Olivier W.,
Steinman R.M.: Quantization of surface antigens on cultured murine epidermal Langerhans
cells: Rapid and selective increase in the level of surface MHC products. J. Invest.
Dermatol., 90: 387-94, 1988.
Parenteau N.L., Nolte C.M., Bilbo P.,
Rosenberg M., Wilkins L.M., Johnson E.W., Watson S., Mason V.S., Bell E.: Epidermis
generated in vitro: Practical considerations and applications. J. Cell Biochem., 45:
245-51, 1991.
Katz S.L: The skin as immunologic organ. J.
Am. Acad. Dermatol., 13: 530-6, 1985.
Thivolet J. Faure M., Demidem A., Mauduit
G.: Long-term survival and immunological tolerance of human epidermal allografts produced
in culture. Transplantation, 42: 274-80, 1986.
Faure M., Mauduit G., Schmitt D., Kanitakis
J., Demidem A., Thivolet J.: Growth and differentiation of human epidermal cultures used
as auto- and allografts in humans. Br. J. Dermatol., 116: 161-70, 1987.>
Swope V.B., Supp A.P., Cornelius J.R.,
Babcock G.F., Boyce S.T.: Regulation of pigmentation in cultured skin substitutes by
cytometric sorting of melanocytes and keratinocytes. J. Invest. Dennatol., 109: 289-95,
1997.
Moseley R.P., Brown J.L, Auld J. et al.: An
immunochemical study of MHC class 1 expression on human Langerhans cells and melanocytes.
J. Pathol., 181: 4, 1997.
Morhenn V.B., Nickoloff B.J., Merigan T.C.
et al.: The effect of gamma interferon on cultured human keratinocytes. J. Invest.
Dennatol., 82: 410, 1984.
Roberts L.K., Krueger G.G., Daynes R.A.: A
functional role for Ia expression by keratinocytes. J. Invest. Dermatol., 82: 418, 1984.
Brain A., Purkis P., Coates P., Hackett M.,
Navsaria H., Leigh I.: Survival of cultured allogenic keratinocytes transplanted to deep
dermal bed assessed with probe specific for Y chromosome. BMJ, 298: 917-9, 1989.
Burt A.M., Pallet C.D., Sloane J.P. et al.:
Survival of cultured allograft in patients with burns assessed with probe specific for Y
chromosome. BMJ, 298: 915-7, 1989.
Cairns B.A., de Serres S., Peterson H.D.,
Meyer A.A.: Skin replacements. The, biotechnological quest for optimal wound closure.
Arch. Surg., 128: 1246-52, 1993.
Gielen V., Faure M,. Mauduit G., Thivolet
J.: Progressive replacement of human cultured epithelial allografts by recipient cells as
evidenced by HLA class 1 antigens expression. Dermatologica, 175: 166-70, 1987.
Acknowledgements. We wish to thank Dr J. Pinies for
generously providing us with the W6/32 anti-HLA A,B,C.
The murine 3T3 Swiss fibroblasts which were used to
initiate this study were a gift from Drs V. Mirabet and A.
Soler. This study was supported by grants from FIS
(Fondo de Investigaciones Sanitarias) and the Basque
Health Service (Ref. 93/0887).
This paper was received on 24 February 2000.Address correspondence to:
Dr Esther Garcia Fernandez,
Dept. de Inmulogia, Hospital de Cruces, Plaza de Cruces
s/n, 48903 Baracaldo, Vizcaya, Spain. Fax: 34 94 600 76;
e-mail: mdgarcia@hcru.osatidetza.net |
|